解碼人胎兒肝臟造血功能
- 2020 年 3 月 27 日
- 筆記
當你的才華還撐不起你的野心時,請潛下心來,腳踏實地,跟著我們慢慢進步。不知不覺在單細胞轉錄組領域做知識分析也快兩年了,通過文獻速遞這個欄目很幸運聚集了一些小夥伴攜手共進,一起成長。
文獻速遞欄目通過簡短介紹,擴充知識面,每天關注,希望你也能有所收穫!
文章資訊
今天分享的這篇文章於2019年10月9日發表在期刊Nature上,文章題目為:Decoding human fetal liverhaematopoiesis 。DOI號為:https://doi.org/10.1038/s41586-019-1652-y.

摘 要
通過10x Genomics 和 Smart-seq2單細胞測序平台對不同胎齡的14個胎肝樣本、7個皮膚樣本、3個腎臟樣本、3個卵黃囊樣本進行測序,獲得了138,575個胎兒肝臟細胞及約74,000個皮膚,腎臟和卵黃囊細胞的單細胞轉錄組譜。推斷了造血幹細胞/多能祖細胞(HSC/MPP)的發育軌跡,定義胎肝造血過程中紅系、淋系及髓系各個階段的造血細胞,首次發現了胎兒皮膚中存在成紅前體細胞,卵黃囊中存在肥大細胞,NK細胞及固有淋巴細胞前體細胞。隨著妊娠的進展,胎肝HSC/MPP分化成為紅細胞的比例降低,髓系及淋系的比例增高,與此同時HSC/MPP的分化潛能也發生了變化。
方法及分析概覽
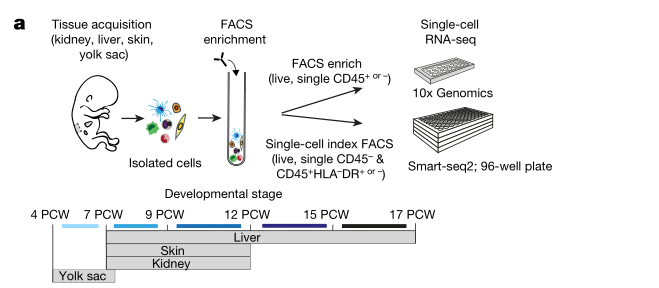
- 樣品:共收集17個胎兒標本,包括14個肝臟樣本、7個皮膚樣本、3個腎臟樣本、3個卵黃囊標本,其中皮膚、腎臟被稱為非淋巴組織(non-lymphoid tissues,NLTs)。樣品詳細資訊如下:

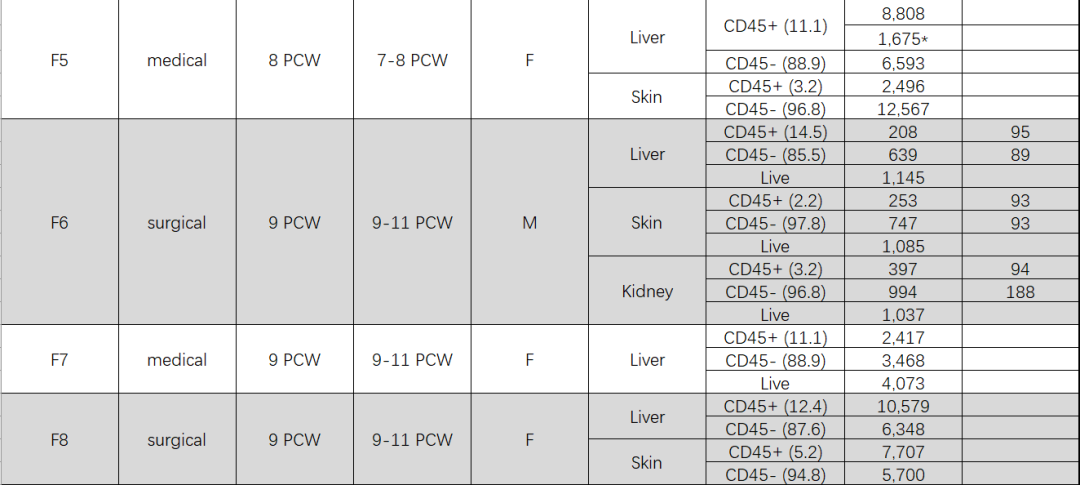

- 建庫測序:10x Genomics樣本通過3』 v2 試劑及 V(D)J Reagent Kits建庫後通過Illumina HiSeq 4000 測序;Smart-seq2樣本通過Illumina Nextera XT kit (Illumina)測序。
- 數據上游分析:10x Genomics 測序數據通過Cell Ranger軟體進行的比對及定量,參考基因組為GRCh38 人類參考基因組。Smart-seq2 測序數據通過STAR軟體進行比對。使用htseq-count(v.0.10.0)計算基因的read計數。
- 品質控制:除去表達< 200個基因且線粒體基因>20%的細胞,除去少於3個細胞中表達的基因。用Scrublet去除Doublets。最終獲得了138,575(n = 14)個胎兒肝臟細胞(使用Smart-seq2分析了1,206個細胞),54,690(n = 7)個皮膚,9,643個腎臟(n = 3)和10,071個卵黃囊(n = 3)細胞。10x Genomics平均每個細胞檢測到約3,000個基因,使用Smart-seq2平均檢測到約6,000個基因/細胞。
- 降維、聚類及細胞定義:應用Seurat包(v2.3.4)進行數據標準化,scaledata, variable gene detection,PCA、TSNE、UMAP降維,Louvain graph-based 聚類。細胞定義通過經典marker及DEG。胎肝細胞聚類又加用了兩種聚類方法:agglomerative clustering及Gaussian Mixture,然後用Rand指數衡量三種聚類方法之間的一致性。細胞定義完後,通過胎肝數據訓練support vector machine (SVM)來構建細胞類型分類器,之後用SVM細胞分類器來定義smart-seq2數據的細胞類型。
- 樣本整合:相同組織樣本之間整合用Harmony,不同組織樣本之間的整合用Merge (Seurat函數)。kBET衡量批次效應。
- 下游分析:通過比較FDG,PAGA和diffusion-map推測發育軌跡。使用Monocle2中的DifferentialGeneTest函數計算偽時變化的基因。CellPhoneDB推斷細胞之間相互作用。
- 功能驗證:免疫組化、免疫熒光、HSC/MPP 培養。
主要結果
1
1.構建了胎兒肝細胞圖譜
- Fig. 1a 實驗流程
- Fig. 1b 胎兒肝臟細胞圖譜
- Fig. 1c 各妊娠階段中每個譜系細胞的比例,胎肝主要以紅系造血為主,但隨著胎齡的增加,紅系比例下降,髓系及淋系比例增加。

1
2. 細胞類型鑒定
- Fig. 2a 展示胎肝各種細胞類型的marker,*表示後續用於流式分選細胞鑒定的marker。
- Fig. 2b 通過選定的marker進行流式分選後,Giemsa染色鑒定細胞類型。

1
3. 胎肝及非淋巴組織的造血
- Fig. 3a FDG圖展示所有的造血細胞,可看到從HSC/MPP向三系的分化軌跡。

- Extended Data Fig. 3a 巨核細胞、肥大細胞、成紅細胞有共同的前體細胞MEMP (megakaryocyte–erythroid–mast cell progenitor)。分化過程中動態調節的基因不同,紅系譜系中的TAL1和KLF1,巨核細胞譜系中的F11R,PBX1和MEIS1,肥大細胞分化中的HES1。

- Fig. 3b 從卵黃囊到肝臟,紅細胞血紅蛋白基因表達以及從原始血紅蛋白HBZ和HBE1表達到胎兒血紅蛋白(HBA1和HBG2)表達的時間變化。

- Extended Data Fig. 3e 肥大細胞,巨核細胞和紅系譜系細胞在肝、卵黃囊、皮膚、腎臟中均顯示出較高的關聯性。

- Extended Data Fig. 3d 皮膚MEMPs表達了一些早期的成紅細胞基因,包括MYL4,表明這些細胞可能充當皮膚中的紅細胞祖細胞,具有造血功能。

1
4. 胎肝和非淋巴組織的淋系細胞生成
- Fig. 4a & 4b 胎肝及非淋巴組織淋系細胞展示。在胎肝中發現了淋系兩個方向分支,NK-T-ILC及B系。與肝臟和卵黃囊相比,非淋巴組織中存在Pro-B,pre-B和B細胞,但缺少HSC / MPP和pre-B前體細胞。

- Extended Data Fig. 5h & 5i NLT中的早期淋巴/ T淋巴細胞,NK細胞和ILC與肝臟相比相同的基因表達趨勢。然而,趨化因子(XCLI和CXCL8)和細胞毒性顆粒(GNLY)基因的組織特異性表達表明,在皮膚和腎臟中發生了NK細胞的成熟和組織適應性。NLT中的ILC前體細胞缺乏其成熟後代ILC1,ILC2和ILC3的全套特徵標記和轉錄因子。

- Extended Data Fig. 5a 根據PAGA, NK細胞(表達NCAM1,CD7,IL2RB和CD3E)和ILC前體(表達KIT,KLRB1,IL7R和RORC)在淋巴分支中具有共同的起源。

- Extended Data Fig. 5e HSC / MPP到B細胞分化過程中動態變化的基因包括SPIB,SP100和CTSS。

1
5. 髓樣細胞組織分布特徵
- Fig. 5a & Fig. 5b 胎兒肝臟,胎盤和卵黃囊的HSC / MPP,髓系祖細胞,單核細胞和巨噬細胞的FDG圖。

- Extended Data Fig. 6a & 6b. DC分化涉及CLEC11A,BATF3和ID2的動態調節,而單核細胞分化涉及S100A8 / A9,FCGR1A / 2A和S100A12的動態調節。漿細胞樣DC(pDC)前體從早期髓樣前體和pre-B前體細胞分化而來,與最近在小鼠中相關研究結果一致。


1
6. HSC/MPP分化潛能隨胎齡變化
- Fig. 6a & 6b FDG展示各個階段的HSC/MPP。根部的HSC表達CLEC9A, HLA-DRA及最高水平的原始基因如MLLT3,為多能的LT-HSC。
- Fig.6c-e 來自CD34 + CD38-CD45RA- HSC / MPP的單細胞培養產生單能和多能的細胞群體。含有紅系細胞的細胞群明顯減少,而含有NK細胞和B細胞的細胞群則隨著胎齡的增加而增加。胎齡小於9周時,無含B細胞的群體產生,提示這一階段B細胞在胎肝中尚未分化。
- Fig 6f-6g胎肝HSC / MPP G0期的比例隨著胎齡的增加而增加,表明HSC逐漸趨於沉默狀態。與臍帶血及骨髓相比,胎肝HSC / MPPs顯示編碼熱休克蛋白(HSPA1A)的基因表達較高,可能用於維持基因組和蛋白質組完整性,而MHC-1(HLA-B)含量較低,表明與抗原呈遞能力較低。

總結
文章從單細胞水平,較為全面的構造了胎兒肝臟造血圖譜,不僅推測出HSC/MPP向三系的分化軌跡,同時發現了HSC/MPP內部的細胞異質性及成熟度的變化,為研究人類其他器官及組織的發育軌跡提供了理想模板。


